|
|
||
Taxon Attribute Profiles
Eucalyptus camaldulensis Dehnh.River Red Gum
IntroductionEucalyptus camaldulensis is a common and widespread tree along watercourses over much of mainland Australia. It is frequently a dominant component of riparian communities, and is an iconic and important species of the Murray-Darling catchment, both ecologically and economically. Taxonomy and EcologyClassificationFamily: Myrtaceae Genus: Eucalyptus- c. 800 species, with all but three or four endemic to Australia. Subgenus: Symphyomyrtus Notes: Eucalyptus camaldulensis exhibits considerable morphological variation throughout its range, and consequently a number of infraspecific taxa have been described. Var. camaldulensis is the most widespread, and the only one occurring in the Murray-Darling Basin. For further discussion on morphological variation, see Brooker et al. (2002). Chemical and genetic variation has also been recorded in E. camaldulensis (e.g. see Doran and Brophy, 1990; Stone and Bacon, 1994; Butcher et al., 2001). Life formEucalyptus camaldulensis is a perennial, single-stemmed, large-boled, medium-sized to tall tree to 30 m high (Bren and Gibbs, 1986), although some authors (e.g. Boland, 1984; Brooker et al., 2002) record trees to 45 m. According to Jacobs (1955) river red gum could reach ages of 500 to 1000 years. See Brooker et al. (2002) for further descriptive information.
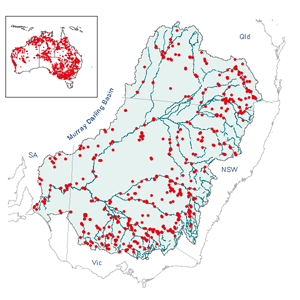
DistributionEucalyptus camaldulensis is found over most of the Australian mainland, except southern Western Australia, south-western South Australia and the eastern coastal areas of Queensland, New South Wales and Victoria (Chippendale, 1988). It is widespread along rivers of all continental Australia (Brooker and Slee, 1996). The accompanying map shows distribution within Australia as well as in the Murray-Darling Basin. HabitatEucalyptus camaldulensis commonly grows on riverine sites, whether of permanent or seasonal water (Brooker et al., 2002). It is most extensive on grey heavy clay soils along river banks and on floodplains subject to frequent or periodic flooding, preferring deep moist subsoils with clay content (Costermans, 1989). It also lines the channels of sandy watercourses and creeks (Boland, 1984), commonly forming ribbon stands but sometimes extending over extensive areas of regularly flooded flats. It can also occur in the higher reaches of creeks in major valleys of hilly country (Cunningham et al., 1981) and infrequently on the margins of salt lakes (CAB International, 2000). In the Murray region it is most commonly found on brown and red clays (Dalton, 1990), and in the Chowilla area it is found along the main Murray River channel and along the backwaters and billabongs (Roberts and Ludwig 1990, 1991). "Status" in communityEucalyptus camaldulensis is generally dominant in the community, commonly forming pure open forests or woodlands (Costermans, 1989). On lower levels of the floodplain, it is usually the only tree species present. On higher areas, it may occur in association with black box (Eucalyptus largiflorens) in the south or coolibah (E. microtheca) in the north (Dalton, 1990). Associated speciesEucalyptus camaldulensis is recorded as occurring with a variety of other tree, shrub and herb species throughout its extensive range, and these are not considered further in this profile. At Chowilla, Roberts and Ludwig (1990, 1991) recorded E. camaldulensis as a dominant species of two riparian communities: "River red gum and reed community" (E. camaldulensis primarily with Phragmites australis), and "River red gum and sedge-rush community" (E. camaldulensis primarily with mixtures of Eleocharis, Juncus, Cyperus and Cynodon dactylon) At Chowilla, E. camaldulensis was recorded in three main communities in a survey undertaken during 1988-1989 (see O’Malley and Sheldon, 1990).
Qualitative and quantitative data – abundance, cover, biomassNo specific data are available relating to cover, abundance or biomass. As noted above E. camaldulensis is a dominant tree in the landscape. Species – interactions with other biodiversityRiver red gum forest wetlands provide habitat for fish and waterbirds (breeding, feeding and refuge areas). This requires a certain length of flooding duration and time of year. Hollows and spouts in river red gum provide habitat for water and forest birds, including two rare species of parrot (Superb Parrot (Polytelis swainsonii) and Regent parrot (Polytelis anthopeplus)) in the Murray River region (Dalton, 1990). Forty-nine phytophagous insects were collected from E. camaldulensis canopies at Gulpa Island State Forest in 1991 and 1992 (Stone and Bacon, 1994). High levels of defoliation have been observed during outbreaks of Uraba lugens (gumleaf skeletoniser) (Dalton, 1990) and Doratifera spp. (cup moths). Mistletoe infestations tend to be localised and occur in stands already stressed by drought or insect attack. Tree death usually only occurs in severe cases (Dalton, 1990). Physiological traits and adaptationsTrees possess deep sinker roots, hypothesised to grow down towards zones of higher water supply (Bren et al., 1991). These roots have extremely high rates of hydraulic conductivity, making them very effective in conducting water (Heinrich, 1990). Seedlings can develop aerenchymatous roots to cope with immersion (see Juvenile period and seedling survival below). It has been suggested (Chesterfield et al., 1984; Chesterfield, 1986 cited in McEvoy, 1992) that the relatively low species richness underneath E. camaldulensis stands in the Barmah forest may be a result of allelopathic suppression from the overstorey. However, others suggest it may be a result of flooding regimes and water stress (see McEvoy, 1992). Leaf shedding reduces water demand by reducing leaf area. It also reduces heat load under dry conditions when transpiration is reduced (Gibson et al., 1994 in Roberts, 2001). Reproduction and Establishment
ReproductionBreeding systemThe eucalypt breeding system is one of mixed mating with preferential outcrossing. Although eucalypts are commonly self-compatible, self-pollination generally results in a reduction in capsule production, seed yield and seedling vigour (see House, 1997). Analyses of the breeding system of E. camaldulensis indicate a predominantly outcrossing mating system (CAB International, 2000). PollinationPollination is mainly by insects but also by birds and small mammals (CAB International, 2000). FloweringEucalyptus flowers in most years from late spring to mid-summer (July to February according to Brooker and Kleinig, 1999, December to February according to Boland, 1984). Flowering intensity is variable and unpredictable from year to year. About 45% of flowers fail to mature (Dexter, 1978). Fruit/seedFruit development and maturation time can be as short as four months (CAB International, 2000). Number of viable seeds per unit weight of a seedlot : mean 698,000/kg (http://www.florabank.org.au/support/articles/sowingtheseeds.doc). Eucalyptus species store little or none of their seed in the soil (see McEvoy, 1992).
EstablishmentEucalyptus camaldulensis is a free producer of seed. Dense stands of young plants appear over extensive areas after floods, at times forming impenetrable thickets. These saplings gradually thin out as they grow (Cunningham et al., 1981). See section on Juvenile period below for more information. In Eucalyptus species, passive release of seed is aided by wind (House, 1997). Free seed fall is least during winter and greatest in spring and summer. High seed fall in spring may have adaptive significance as floods usually recede during this period (Dexter, 1978). Eucalyptus camaldulensis seeds sank within 36 hours of being dropped into still water in laboratory tests and it was suggested that under field conditions they would sink more rapidly (Dexter, 1978). However, McEvoy (1992) found that seeds remained buoyant for at least 17 days after sowing. He suggested that there might be a potential for floodwaters to act as a dispersal agent. Dispersability; establishment and growth
Saplings gradually thin out as they grow, to form forests of straight-trunked trees. In more arid regions, where ribbon stands occur along creeks, the tree is more gnarled and develops a large spreading canopy. This form also occurs throughout the region wherever the tree grows in isolation on deep fertile soils with a good moisture supply. Eucalyptus camaldulensis has probably one of the fastest growth rates for a tree and with a good water supply can attain a height of 12-15 m in a few years (Cunningham et al., 1981). Competition for moisture by ground vegetation and /or overstorey trees can influence seedling survival depending on seasonal conditions and flooding. The availability of moisture is greatly reduced within the zone of influence of trees (which may extend to 40 m around a mature tree). In the absence of competition seedling survival is 20-30 times greater (Dexter, 1978). Juvenile periodGeneration time may be as short as three years from planting to the production of the first seed crops (CAB International, 2000). Precocious flowering may occur as early as six months (Khan, 1965, cited in House, 1997). For wild trees the time to first flowering is more likely to be five years for a few scattered individuals and 7-10 years for general flowering. The seed from early flowerings is usually very disappointing in terms of germination capacity and seedling growth, probably reflecting high levels of inbreeding (pers. comm., J. Doran, 2004). Fruit development and maturation time can be as short as four months (pers. comm., J. Doran, 2004). Seedling establishment rather than germination is the critical stage in stand regeneration. Flood timing affects germination success. Flood recession in spring-early summer is optimal for regeneration while winter floods with winter recession are unfavourable. Spring-summer floods followed by summer recession provide suitable germination conditions but subsequent heat and water stress can cause massive seedling mortality. Germination can happen without flooding if the winter is wet. If seedlings survive frost, but conditions continue dry, moisture stress in the following summer is likely (Roberts and Marston, 2000). Seedlings are vulnerable during the establishment phase to heat stress and immersion. Seedlings cope with heat stress by developing roots giving good penetration into the sub-soil and accessing soil moisture. Seedlings also develop resilience early, allowing them to shed leaves in times of moisture stress and recover from axillary buds when moisture is again available (Dexter, 1978). Seedlings develop adventitious roots and aerenchymatous tissue to deal with anoxia resulting from immersion (Heinrich, 1990). Complete immersion, unless brief, is likely to kill seedlings; lower leaves of small saplings die if submerged for long periods (Roberts and Marston, 2000). Seedlings increase tolerance to flooding with age. Two-month old seedlings can survive waterlogging for one month (Marcar, 1993), while seedlings 50-60 cm tall can survive extended flooding of 4-6 months and complete immersion for a few weeks by shedding leaves (Dexter, 1978). The ability of the species to compete with weeds is poor when young (pers. comm., J. Doran, 2004). Hydrology and salinityHydrologyEucalyptus camaldulensis obtains its water from three main sources: ground water, rainfall and river flooding. It is river flooding which enables the species to survive in semi-arid areas. The unregulated flooding regime in western New South Wales consisted of peak flows in late winter and spring with low flows in summer and autumn (Dalton, 1990). Changes in the river flow patterns of the Murray, as a result of large scale dam building, has led to reduced extent and depth of winter flooding, reduced frequency of flooding, increased duration of non-flood periods, increased occurrence and variability of summer floods, increased river flow capacity (as a result of desnagging) and decreased total annual flow. These changes have produced major deterioration in much of the riparian forest, including reduced tree growth rate, accelerated mortality and minimal regeneration (Bacon et al., 1993). Stands of river red gum are intimately associated with the surface-flooding regime of the watercourses and related ground water flow. The high water use of river red gums contributes to maintaining watertables at depth (Dalton, 1990). Stabilised water levels are characteristic of large parts of the Chowilla floodplain, (Roberts and Ludwig, 1991). At the time of the Chowilla floodplain biological study (O'Malley and Sheldon, 1990) there was a high incidence of dieback amongst river red gum and black box woodland associations. It appears that woodland areas removed from the main channel or anabranch creeks are more susceptible to dieback. Dieback is variously attributed to altered hydrologic regime (reduced frequency and depth of floodplain inundation) or increasingly saline soils (due to mobilisation of saline groundwater as a consequence of the hydraulic pressure exerted by the lock). At Chowilla the two riparian communities described by Roberts and Ludwig (1990) were found in two distinct places. The river red gum and sedge-rush community occurred in riparian habitats where current was slow and the bank was gently sloping and not subject to strong wave action. The river red gum and reed community was associated with relatively fast currents and steep banks exposed to strong wave action. Regeneration of river red gum was recorded at several channel edge localities, especially where the channel bank was not far elevated from the anabranch creek level (O'Malley and Sheldon, 1990). Before the introduction of regulation on the Murray River, groundwater in the Chowilla region flowed under the floodplain into the river. After the installation of locks, which also resulted in the previously ephemeral creeks being continuously filled with water, the direction of groundwater flow was reversed. The groundwater now discharges into the anabranch creeks and into the soil (Jolly and Walker, 1995). Most recharge of the groundwater system at Chowilla is dominated by flooding. The heavy clay soils in the area also act to decrease the impact of rainfall. Three types of flood recharge occur. ‘Bank recharge’ is when a stream recharges the aquifer through the bank. ‘Diffuse recharge’ is the recharge of the groundwater through the soil surface after the stream has broken its banks. In ‘localised recharge’ the floodwater infiltrates through isolated areas of the floodplain at a higher rate (e.g. in old depressions, dunes with a thin clay layer or old meanders). See Jolly and Walker (1995) for a discussion on the different impacts of the three types of recharge.
Salinity toleranceEucalyptus camaldulensis demonstrates moderate salt tolerance (Benyon et al., 1999). Benyon et al. (1999) showed that increasing salinity is associated with reduced tree growth in an experiment on a saline discharge site near Wellington, NSW. Growth was better for E. camaldulensis trees planted on non-saline soil than on moderately saline soil. An increase in soil salinity was associated with a decrease in the average leaf area per tree. Thorburn et al. (1994) showed that river red gums in the Chowilla floodplain were not obtaining all their water from the creek, even when the trees were over highly saline groundwater. Eucalyptus camaldulensis trees at Chowilla that only had access to surface water during a flood were not utilising low-salinity floodwaters in preference to more saline groundwater during a flood period (Thorburn and Walker, 1994). Results indicated that the trees might be less affected by changes in creek flow and/or salinity than was previously thought (also see Thorburn et al., 1992; Mensforth et al., 1994). Flooding regimesThe change in the river flow has led to a decline in river red gum health and changes in the understorey composition. Permanent inundation leads to river red gum death (Dalton, 1990). However, river red gums have survived relatively long periods of continuous flooding (24 months at Barmah and 3-4 years behind the Hay Weir (Bren, 1987)). Field observations suggest river red gums can survive 2-4 years of continuous flooding before showing signs of stress (Roberts and Marston, 2000). Forest flooding, particularly in late winter, is a key factor in controlling the leaf skeletoniser moth by: providing conditions favourable to the growth of a fungal pathogen of the insect (Aspergillus); removing pupation sites within the ground litter; drowning the insect larvae. Reduction of flooding frequency through regulation has advantaged these insects (Dalton, 1990). Change in water regimesReduced flooding has resulted in less water being available for regeneration and seasonal growth. Permanent inundation results in tree death. Other vegetation communities have adapted to infrequent flooding and are able to expand, usually at the expense of river red gum communities (Dalton, 1990). Eucalyptus camaldulensis was seen to be ‘invading’ a natural grassland in the Barmah-Millewa Forest, presumably as a result of changes in river regulation (Bren, 1992) Response to disturbance (non-hydrological)GrazingRabbits and kangaroos heavily graze seedlings during prolonged dry periods when feed is scarce (Dexter, 1978). Until 1950s grazing of river red gum forests was at a fairly high level, modifying the original understorey (Dalton, 1990). Where narrow bands of trees occur along a watercourse, too high grazing pressure will disadvantage maintenance of a self-replacing stand. In these cases only a small amount of regeneration results, and this is easily grazed out by stock. However, sapling growth is not, or rarely, grazed by stock unless animals are starved of other forage (Cunningham et al., 1981). Cattle grazing on weeds may help control weeds, reducing competition for moisture (Dexter, 1978). FireEucalyptus camaldulensis is very fire sensitive and even low intensity fires may cause cambial injury (Dexter, 1978). Fire kills regeneration and even mature trees are susceptible if the fire is intense enough since E. camaldulensis lacks a lignotuber. Fire will cause damage to the butt, lowering the value of the timber and predisposing tree to fungal and insect attack (Dalton, 1990). OtherFeral pigs can disturb large areas through digging and wallowing, causing erosion and destroying wetland areas (Dalton, 1990). Conservation statusEucalyptus camaldulensis is one of the most widespread tree species across Australia, and is not considered at risk.
Uses (including ethnobotanical)River red gum forests are historically and culturally important due to the number of significant Aboriginal sites they contain. Common relics include canoe and shield trees. Such trees show scars where the bark was removed (Dalton, 1990). The wood has been used for heavy construction, railway sleepers, flooring, framing, fencing, plywood and veneer manufacture, wood turning, firewood and charcoal production (Boland, 1984). Eucalyptus camaldulensis is of major importance in Australia as a source of honey, producing heavy yields of nectar in good seasons (Clemson, 1985). It also provides bees with an important source of good quality pollen (CAB International, 2004). Due to its natural adaptation to both temperate and tropical climates with both winter and summer rains, river red gum is the most widely planted species in arid and semi-arid regions around the world, primarily in timber plantations (Eldridge et al., 1993 in CAB International , 2000). SummaryCompared with most species, there is a considerable bank of knowledge relating to Eucalyptus camaldulensis and its functioning in the landscape, and in particular its performance in the Murray-Darling Basin. From past changes in water regimes we know that E. camaldulensis is affected by changing water levels and that mature stands have been lost through permanent flooding. Flood timing affects germination success, and seedling establishment, the critical stage in regeneration, is vulnerable to heat stress and immersion. Eucalyptus camaldulensis is not physiologically adapted to either drought or salinity, although these stresses can be tolerated for short periods or at low levels. Stands of river red gum are associated with the surface flooding regime of watercourses and related ground water flow. The species is a profligate and opportunistic water user, and this is a contributing factor to the maintenance of water tables at depth. Even without large amounts of empirical data it is clear that loss of large tracts of the species in the Murray River corridor would have a major impact on the hydrology of the system, as well as on vegetation communities and associated biodiversity. ReferencesBacon, P.E., Stone, C., Binns, D.L., Leslie, D.J. and Edwards, D.W. (1993) Relationships between water availability and Eucalyptus camaldulensis growth in a riparian forest. Journal of Hydrology 150, 541-561. Benyon, R.G., Marcar, N.E., Crawford, D.F. and Nicholson, A.T. (1999) Growth and water use of Eucalyptus camaldulensis and E. occidentalis on a saline discharge site near Wellington, NSW, Australia. Agricultural Water Management 39, 229-244. Boland, D.J., Brooker, M.I.H., Chippendale, G.M., Hall, N., Hyland, B.P.M., Johnston, R.D., Kleinig, D.A. and Turner, J.D. (1984) Forest Trees of Australia. Nelson and CSIRO, Melbourne. Bren, L. (1990) Red Gum Forests. In Mackay N. and Eastburn, D. (eds) The Murray. Murray-Darling Basin Commission, Canberra, 230-242. Bren, L.J. (1987) The duration of inundation in a flooding river red gum forest. Australian Forest Research 17, 191-202. Bren, L.J. (1992) Tree invasion of an intermittent wetland in relation to changes in the flooding frequency of the River Murray, Australia. Australian Journal of Ecology 17, 395-408. Bren, L.J. and Gibbs, N.L. (1986) Relationships between flood frequency, vegetation and topography in a river red gum forest. Australian Forest Research 16, 357-370. Brooker, M.I.H. and Kleinig, D.A. (1999) Field Guide to Eucalypts, South-eastern Australia. Volume 1, Bloomings Books, Hawthorn. Brooker, M.I.H. and Slee, A.V. (1996) Eucalyptus. In Walsh, N.G. and Entwisle, T.J. (eds) Flora of Victoria, Volume 3, Dicotyledons: Winteraceae to Myrtaceae, Inkata Press, Melbourne. Brooker, M.I.H., Connors, J.R., Slee, A.V. and Duffy, S. (2002) EUCLID: eucalypts of southern Australia (CD Rom), CSIRO Publishing, Collingwood. Butcher, P.A., Otero, A., McDonald, M.W. and Moran G.F. (2001) Nuclear RFLP variation in Eucalyptus camaldulensis Dehnh. from northern Australia. Heredity 88: 402-412. CAB International. (2000) Eucalyptus camaldulensis. Forestry Compendium Global Module. CAB International, Wallingford, UK. Chesterfield, E.A. (1986) Changes in the vegetation of the river red gum forest at Barmah, Victoria. Australian Forestry 49, 4-15. Chippendale, G.M. (1988) Flora of Australia, Volume 19, Myrtaceae, Eucalyptus, Angophora. Australian Government Publishing Services, Canberra. Costermans, L. F. (1989) Native trees and shrubs of south-eastern Australia,Weldon, Sydney. Cunningham, G.M., W.E.Mulham, P.E.Milthorpe and J.H.Leigh (1981) Plants of Western New South Wales, Soil Conservation Service of New South Wales. Dalton, K. (1990) Managing our river red gums. Soil Conservation Service of New South Wales, Sydney. Dexter, B.D. (1978) Silviculture of the River Red Gum forests of the central Murray floodplain. Proceedings of the Royal Society of Victoria 90, 175-194. Doran, J. and Brophy, J.J. (1990) Tropical gums – a source of 1,8-cineole-rich Eucalyptus oil. New Forests 4, 157-178. Heinrich, P. (1990) The eco-physiology of riparian River Red Gum (Eucalyptus camaldulensis) Final Report, Australian Water Resources Advisory Council. House, S.M. (1997) Reproductive Biology of Eucalypts in Williams, J. and Woniarski, J. (eds). Eucalypt ecology: individuals to ecosystems. Cambridge University Press. Jacobs, M.R. (1955) Growth Habits of the Eucalypts. Forestry and Timber Bureau, Canberra Jessop, J.P. (1986) Myrtaceae. In Jessop, J.P. and Toelken, H.R., Flora of South Australia: Part II, Leguminosae – Rubiaceae, South Australian Government Printing Division, Adelaide. Jolly, I.D. and Walker, G.R. (1995) A sketch of salt and water movement in the Chowilla floodplain, CSIRO Division of Water Resources. McEvoy, P.K. (1992) Ecophysiology of 3 Eucalyptus species on the River Murray floodplain. Unpublished thesis, M.For Sci, Univeristy of Melbourne. Mensforth, L.S., Thorburn, P.J., Tyerman, S.D. and Walker, G.L. (1994) Sources of water used by riparian Eucalyptus camaldulensis overlying highly saline groundwater, Oecologia 100, 21-28. O'Malley, C. and Sheldon, F. (1990) Chowilla floodplain biological study. Nature Conservation Society of South Australia, Adelaide Roberts, J. and Ludwig, J.A. (1990) Riparian habitats on the Chowilla floodplain of the River Murray, South Australia, Wetlands (Australia) 9, 13-19 Roberts, J. (2001) Large plants. In Young, W.J.(ed), Rivers as Ecological Systems - the Murray-Darling Basin, pp. 187-221. Murray-Darling Basin Commission, Canberra. Roberts, J. and Marston, F. (2000) Water regime of wetland and floodplain plants in the Murray-Darling Basin. A sourcebook of ecological knowledge., CSIRO Land and Water, Canberra. Stone, C. and Bacon, P.E. (1994) Relationships among moisture stress, insect herbivory, foliar cineole content and the growth of river red gum Eucalyptus camaldulensis. Journal of Applied Ecology 31: 604-612. Thornburn, P., Walker, G. and Hatton, T. (1992) Are river red gums taking water from soil, groundwater or streams? Catchments of green: a national conference on vegetation & water management, conference proceedings - volume B, 63-68, Greening Australia, Canberra. Thorburn, P.J. and Walker, G.R. (1994) Variations in stream water uptake by Eucalyptus camaldulensis with differing access to stream water. Oecologia 100:293-301. |